Research
Unlike multicellular and unicellular organisms, whose units are mostly fixed to a location, a honey bee colony is a superorganism, with moveable units that interact with each other and with their nest environment to gather information.
Researchers in the Smith Lab are encouraged to develop their own research program within the broad framework of social physiology and understanding how honey bee colonies function.
Below are some examples of the type of work we do.
For a full list of lab publications, see here.

Photo: Ben Koger
Lifetime tracking of honey bees
We know that workers of different ages perform different tasks in different parts of the nest. Young bees, for example, perform brood care in the center of the nest, while older bees advertise food sites on the dance floor, near the nest entrance. This system of temporal polyethism means that workers of different ages are located in different parts of the nest. It is unknown, however, how individuals change the way they move to get to the “right place” (nest location) at the “right time” (worker age). One could think of this as the circulatory system of the superorganism.
This type of research was previously unimaginable, because it would take so much time to watch each individual bee. Indeed, much of what we knew about the individual tasks of a honey bee originate from Martin Lindauer’s impressive work on bee #107 (he watched this bee for 7.5+ hrs per day, for 24 days). Instead of watching each bee individually, we use the BeesBook tracking system, developed by our close collaborators, the Landgraf Lab. We use this system to track bees throughout their entire lives, to understand the variability in individual life histories.
Growth, development, and reproductive shifts in honey bee colonies
All living systems have cycles of growth, development, and reproduction. Developmental cycles regulate how organisms invest resources, how they behave, and how they interact with their environment. These cycles, just like the organisms themselves, are under constant selective pressure to allocate resources adaptively in a stochastic biotic and abiotic environment. While this problem is universal, from unicellular yeast, to multicellular mammals, to superorganismal social insects, our understanding of how individual units detect the developmental state of the whole organism is limited.
One such developmental shift is the onset of reproductive investments, akin to puberty. In honey bee colonies, this occurs when workers collectively shift to building reproductive comb, for rearing drones. We know that the workers will only do this once the colony has surpassed a threshold colony size, but how do the workers detect that they are living in a colony that can “afford” to invest in reproduction?
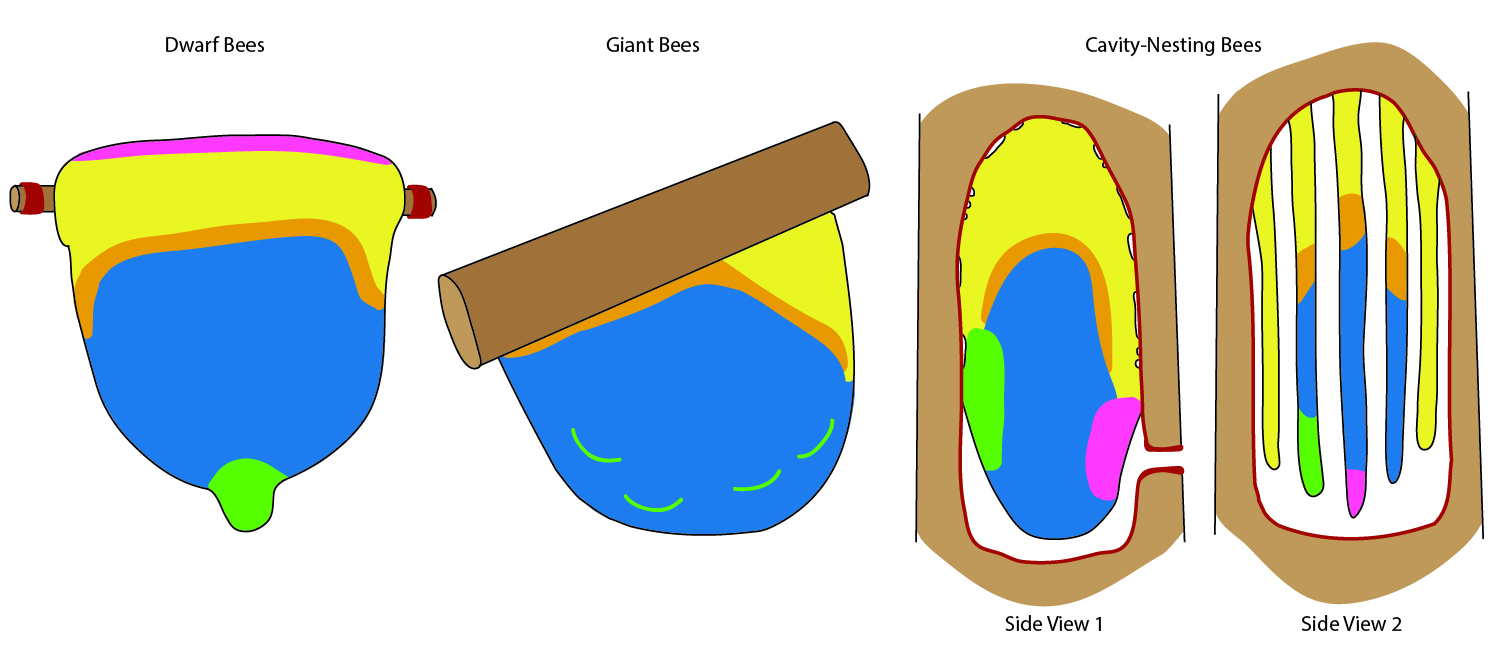
Nest construction, architecture, and organization
Honey bee nests inspire awe at multiple levels–from the perfection of the stacked hexagons, to the organization of the nest by the unsupervised collective. The honey bee nest is tightly linked to colony function. It is where new workers are reared, food is stored, and information is exchanged. The honey bee nest is a prime example of an extended phenotype – an external structure built by an organism that contributes to its fitness. Social insect nests provide extreme examples of the extended phenotype, because the nest itself is critically important for all aspects of colony life. In the same way that one might study the behavior and physiology of a “traditional” organism, we apply the same principles to the nest of the honey bee.